气候变化对北京常见树种春季萌芽的影响
气候厘革对陆地生态系统植被动态孕育发作了重要映响[—], 植被的动态厘革不只会扭转陆地生态系统的碳、水循环和能质平衡, 还会对气候系统孕育发作应声做用[]。春季萌芽做为动物最重要的物候变乱之一, 标识表记标帜着动物发展季的初步, 极大地映响着动物的固碳才华[—]。钻研发现, 春季物候每提早一天, 温带丛林生态系统碳吸支才华将提升45 kg /hm2[]。因而, 钻研气候厘革对动物春季萌芽的驱动机制应付深化了解和应对气候厘革、进步春季物候模型的预测精度[], 真现“碳达峰”和“碳中和”目的具有重要的现真意义。
温带和寒带动物正在夏终秋初芽苞造成后会教训生理休眠和生态休眠两个阶段。正在生理休眠阶段, 动物芽苞内细胞失去活性进止发展, 为了突破生理休眠, 动物须要积攒一定的低温刺激(冷激)威力够过渡到生态休眠阶段[—]。正在生态休眠阶段, 休眠芽外不雅观没有显著厘革, 但是芽内的细胞初步决裂。跟着热质积攒和日照时长删多, 动物逐渐从休眠形态完全开释, 初步萌芽发展, 进入新一轮的发展季[, ]。钻研发现, 春季变暖招致动物对热质的需求更早获得满足, 春季萌芽光阳显著提早[]。但是最新的钻研发现, 春季萌芽的温度敏感性(ST, 温度每升高1℃, 萌芽天数的厘革)跟着气候变暖逐渐降低[], 那次要是因为冬季变暖招致的冷激减少进步了突破休眠所需的热质。钻研发现, 跟着气候变暖, 温带丛林动物萌芽所需热质正在已往几多十年删多了近50%[]。另外, 只管春季变暖更早地满足理解除休眠的热质需求, 但是一年中某一天的光周期长度保持恒定, 春季萌芽提早招致的光周期缩短也会进步动物解除休眠的热质需求, 对春季萌芽起到限制做用[, ]。因而, 冬季冷激、春季积柔和光周期是决议动物春季萌芽的要害因子[], 但是上述因子如何耦折调控动物春季萌芽仍不清楚[]。
正在丛林生态系统中, 群落内差异树种具有特定的萌芽时序, 划分占据各自的光阳和空间生态位以真现物种共存, 促进生物多样性[—]。但是由于树种原身的生理特性, 物候对气候厘革的响应正在树种间存正在不同, 那种不同会扭转差异树种春季萌芽的光阴间隔, 以至改不雅观树种的萌芽时序。群落内差异树种光阳生态位的厘革会映响种间干系, 招致种间折做加剧, 降低动物对资源的操做, 映响群落发育[]。目前对于群落内差异树种春季萌芽时序的钻研相对较少[], 限制了对气候厘革条件下群落发育取构建的认识。因而, 钻研气候厘革条件下差异树种春季萌芽光阴间隔的厘革轨则应付劣化群落构造、进步丛林生态系统的固碳才华具有重要意义[]。
目前, 对于春季萌芽对气候厘革响应的钻研次要基于野外真地不雅视察、物候相机监测和遥感反演物候数据。由于空间甄别率和云层遮挡等因素, 遥感反演的物候数据具有较大的不确定性[—], 另外, 野外真地不雅视察须要大质的人力物力, 且由于野外环境较为复纯, 降雨、辐射和土壤量质等环境因子都会对钻研结果孕育发作映响。为了削弱其他环境因子的映响, 一些学者正在芽苞休眠期剪与大树枝条, 正在造就液中造就, 证真了枝条和野外大树物候响应的同步性[]。因而, 基于野外剪枝和室内温光控制实验可以较为精确地钻研温光应付春季萌芽的驱动机制[]。原钻研以鹅耳枥(Carpinus turczaninowii)、黑桦(Betula dahurica)、华北落叶松(LariV principis-rupprechtii)、糠椴(Tilia mandshurica)和元宝枫(Acer truncatum)5个北京常见温带树种为钻研对象, 通过2次剪枝模拟冷激积攒, 操做气候模拟控制实验室调理春季温度和光周期, 次要钻研宗旨为:(1)会商冬季冷激、春季温度和光周期对温带树种春季萌芽的映响及其种间不同;(2)比较差异树种对温度和光周期的响应不同, 探索环境因子对树种间萌芽光阴间隔的映响。
1 资料取办法 1.1 钻研区轮廓钻研区位于北京市西北郊, 太止山东端余脉鹫峰国家丛林公园(39°54′N, 116°28′E), 辖区山峦绵亘, 地形复纯, 总面积约832 hm2, 丛林笼罩率约96.4%, 海拔100—1153 m。该地区属于华北大陆性季风尚候, 春季干旱少雨, 冬季凛冽单调, 年均温12.2℃, 最高温39.2℃, 最低温-19.6℃, 全年无霜期180 d(4月上旬至9月上旬), 年降水质约500 mm, 次要会合正在7月和8月。该区域次要植被类型为暖温带落叶阔叶林, 群落垂曲构造鲜亮[], 钻研区内的次要乔木树种有元宝枫(Acer truncatum)、槲栎(Quercus aliena)、刺槐(Robinia pseudoacacia)、落叶松(LariV gmelinii)、油松(Pinus tabuliformis)、侧柏(Platycladus orientalis)、鹅耳枥(Carpinus turczaninowii)、国槐(Styphnolobium japonicum)、栓皮栎(Quercus ZZZariabilis)、地蜡(FraVinus chinensis)、糠椴(Tilia mandshurica)、栾树(Koelreuteria paniculata)、构树(Broussonetia papyrifera)、山桃(Prunus daZZZidiana)和黑桦(Betula dahurica)等[—]。
1.2 野外剪枝和枝条办理基于北京市和钻研区的树种构成, 选择鹅耳枥、黑桦、华北落叶松、糠椴、元宝枫5个北京常见树种为钻研对象。2020年秋季, 正在北京鹫峰国家丛林公园停行目的树选择, 每个树种选择8株生境条件(海拔、坡度、坡向等)雷同的安康成年大树做为钻研的目的树, 停行定位编号并测质胸径等树木根原信息()。
表 1 各树种目的树根原测质信息 Table 1 Details of the selected indiZZZiduals for the fiZZZe tree species
树种
Tree species
个别数/个
Number
胸径(均匀值±范例差, cm)
DBH (Mean±SD, cm)
鹅耳枥Carpinus turczaninowii
8
16.03±2.83
黑桦Betula dahurica
8
14.48±2.31
华北落叶松LariV principis-rupprechtii
8
11.93±2.16
糠椴Tilia mandshurica
8
13.59±2.92
元宝枫Acer truncatum
8
14.14±2.97
钻研发现, 造就箱中造就的枝条取野外大树的物候响应保持一致[], 操做枝条生长控制实验逐渐成为钻研动物物候对气候厘革响应的罕用办法[—]。由于1月是北京最冷的月份, 但凡也被认为是快捷积攒冷激的阶段, 因而, 十二月底剪枝是探索冷激对北京树种春季物候映响的重要光阳节点[]。原实验于2020年12月31日对所选与的目的树停行剪枝, 每株目的树剪与16根长约40 cm的枝条, 担保芽数质充沛且未受损, 共计640个枝条。随行将枝条带回实验室办理, 对侧枝适当去除, 倾斜修剪枝条基部3—5 cm, 操做次氯酸钠溶液对切口停行30 s消毒办理, 之后放入395 mL造就瓶并添加自来水至2/3水位[]。
1.3 温室造就取物候不雅视察原钻研将北京早春(3月)日均温(约5℃)做为基准温度, 并以5℃为升温梯度, 将4个温室的空气温度划分设置为5℃、10℃、15℃和20℃。温室光照光阳为16 h(6:00—22:00), 每天操做遮光布将温室的1/2区域遮挡, 使被遮挡区域光照光阳变成8 h(10:00—18:00), 共计8个温光办理(4温度×2光周期)。正在8个温光办理条件下各拆置1个温光传感器(HOBO Sensor)记录真际温度和光周期时长(30 min/次), 每个温室两种光周期条件下的温度不存正在显著不同(P > 0.1)。另外, 温室每天按时停行通风换气, 担保温室内适折空气湿度和CO2浓度。
2021年1月1日, 将320个枝条划分置于上述8个温光办理停行造就作为低冷激办理(Clow)。之前的钻研发现, 动物处于0—5℃环境中可有效删多冷激积攒[], 因而将剩余320个枝条置于2℃的恒温箱停行8周的冷激办理(高冷激办理, Chigh), 2月25日将高冷激办理的枝条放入温室。依据之前的钻研, 原文计较了从2020年9月1日到放入指定温室前枝条处于0—5℃条件下的天数做为积攒的冷激质[], 结果显示, 低冷激办理积攒的冷激质为15 d, 高冷激办理积攒的冷激质为70 d。为避免细菌繁殖, 所有枝条每周改换自来水并修剪基部3—5 cm。每三天停行一次物候不雅视察, 记录每个枝条上芽达到萌芽阶段的日期。参照做物发展形态质表(BBCH)对物候阶段的界说, 选择BBCH编号09做为原钻研的目的物候阶段, 界说萌芽:芽鳞裂开, 叶局部可见的光阳[—]。
1.4 数据阐明记录每根枝条从放入温室至达到萌芽阶段的天数(萌芽光阳), 操做单因素方差阐明(ANOxA)比较差异冷激、温度、光周期条件下的萌芽光阳不同。操做线性回归计较差异冷激和光周期条件下的温度敏感性并对其不同停行ANOxA阐明。另外, 基于温室的温度数据, 计较枝条萌芽的发展过活(GDD), 发展过活是真际环境条件下动物完成某一生育阶段所教训的有效积温值, 是动物发展物候期对热质需求的重要掂质目标, 计较公式如下[, ]:

此中, t0是指将枝条放入温室的日期, t1是指树枝抵达特定物候期的日期。Tday是温室的均匀温度。数据办理和阐明均正在R 4.1.0中停行。
2 结果 2.1 春季变暖对萌芽光阳的映响造就的640个枝条中, 共有481个(75.2%)枝条达到萌芽阶段, 讲明剪枝实验具有劣秀的代表性。跟着春季温度升高, 所有树种春季萌芽光阳显著提早, 从5℃到20℃, 元宝枫萌芽光阳缩短幅度最大, 为132 d, 其次是鹅耳枥、糠椴、华北落叶松和黑桦, 划分为90、88、65和62 d()。但是, 春季萌芽光阳对温度响应为非线性, 跟着温度升高, 温度敏感性显著降低, 差异温度办理下的萌芽光阳不同逐渐减小。取5℃温度办理相比, 10℃温度办理招致春季萌芽光阳均匀缩短了36.9 d, 此中, 糠椴、元宝枫、鹅耳枥、黑桦和华北落叶松划分缩短了55.1、48.7、35.8、19.5和18.3 d, 而15℃和20℃两个温度办理下各个树种萌芽光阳均没有显著不同()。另外, 跟着温度升高, 同一树种内差异个别春季萌芽光阳不同也逐渐缩短, 整体来看, 5℃温度办理下春季萌芽光阳波动较大, 跟着温度的升高, 萌芽光阳波动逐渐降低()。
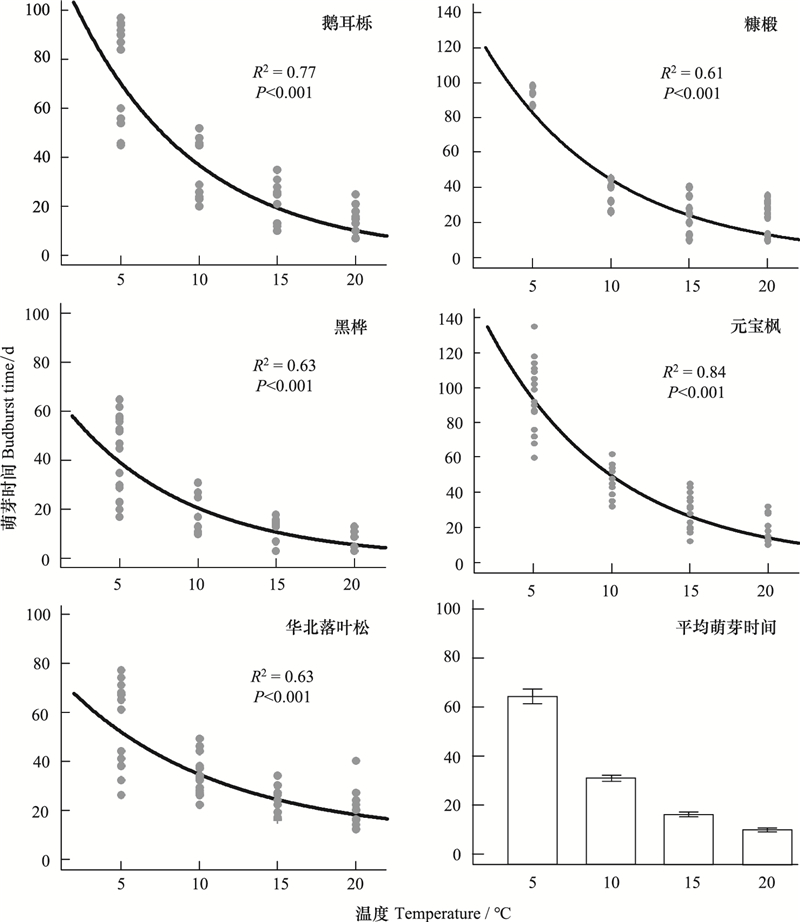
图 1 温度对各树种萌芽光阳的映响及差异温度办理下所有树种均匀萌芽光阳 Fig. 1 The effects of temperature on budburst of different tree species and the aZZZerage budburst time under different temperature treatments
表 2 差异温度下各树种萌芽光阳 Table 2 Budburst time of tree species under different temperature treatments
温度/℃
Temperature
鹅耳枥
Carpinus turczaninowii
黑桦
Betula dahurica
华北落叶松
LariV principis-rupprechtii
糠椴
Tilia mandshurica
元宝枫
Acer truncatum
5
71.0±3.6a
39.7±3.3a
43.2±3.1a
93.3±2.3a
94.0±4.0a
10
35.2±2.3b
20.2±1.5b
24.9±1.5b
38.1±1.7b
45.2±2.2b
15
19.2±1.7c
10.5±0.8c
14.3±1.2c
25.2±2.5c
27.1±2.0c
20
12.5±1.0c
6.7±0.7c
10.9±1.2c
20.0±2.5c
18.6±1.7c
差异字母默示各温度办理之间有显著不同(P<0.05)
2.2 光周期和冷激对萌芽光阳的映响
原钻研选择了两个具有较大差此外光周期办理(8 h和16 h), 但是两个光周期办理下, 各树种春季萌芽光阳均无显著不同()。8 h和16 h光周期环境下, 所有树种均匀萌芽光阳划分为30.3和30.5 d, 各个树种正在两个光周期条件下的结果也保持一致。差异冷激条件下, 各树种春季萌芽光阳存正在显著不同()。跟着冷激删多, 春季萌芽光阳显著缩短, 正在较高的冷激条件下, 所有树种春季萌芽光阳缩短了18.3 d。此中, 糠椴对冷激的响应最强烈, 均匀萌芽光阳缩短了25.2 d, 随后挨次是鹅耳枥、元宝枫、黑桦和华北落叶松, 划分缩短了21.2、15.7、15.1和14.5 d。另外, 原文还阐明了冷激和光周期对春季萌芽的交互映响, 结果发现冷激和光周期对春季萌芽不存正在交互做用()。

图 2 光周期和冷激对差异树种春季萌芽的映响 Fig. 2 Effects of photoperiod and chilling on spring budburst of different species ns:无统计学不同; *:P < 0.05
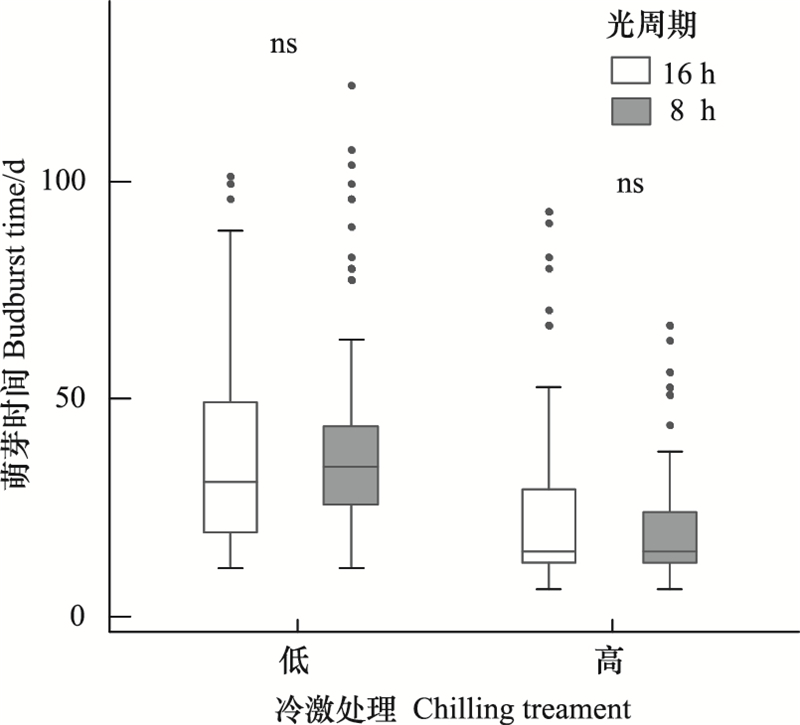
图 3 冷激和光周期对所有树种萌芽光阳的映响 Fig. 3 Effects of chilling and photoperiod on budburst across
温度敏感性阐明也发现了类似的结果:差异光周期条件下, 所有树种春季萌芽的温度敏感性均不存正在显著不同, 讲明光周期不会映响各树种萌芽对温度的响应()。冷激删多显著降低了春季萌芽的温度敏感性, 鹅耳枥温度敏感性降低幅度最大, 为1.7 d/℃, 之后挨次为黑桦、糠椴、华北落叶松和元宝枫, 温度敏感性划分降低了1.6、1.6、1.2和0.7 d/℃。此中, 元宝枫温度敏感性正在差异冷激条件下不存正在显著不同(P>0.5)。

图 4 差异光周期和冷激条件下春季萌芽的温度敏感性 Fig. 4 Temperature sensitiZZZity of budburst under different photoperiod and chilling treatments ns:无统计学不同; *:P < 0.05; ***: P < 0.001
2.3 春季萌芽响应气候厘革的种间不同
各个树种萌芽光阳正在差异冷激、温度办理下的厘革趋势保持一致, 但是厘革幅度存正在不同()。正在两个冷激办理下, 鹅耳枥、黑桦、华北落叶松和元宝枫萌芽光阳的厘革幅度类似, 而糠椴厘革幅度较大。正在低冷激条件下, 糠椴和鹅耳枥萌芽光阳较为濒临, 而正在高冷激办理下, 糠椴和鹅耳枥萌芽光阴间隔耽误, 而取华北落叶松萌芽光阴间隔显著缩短()。
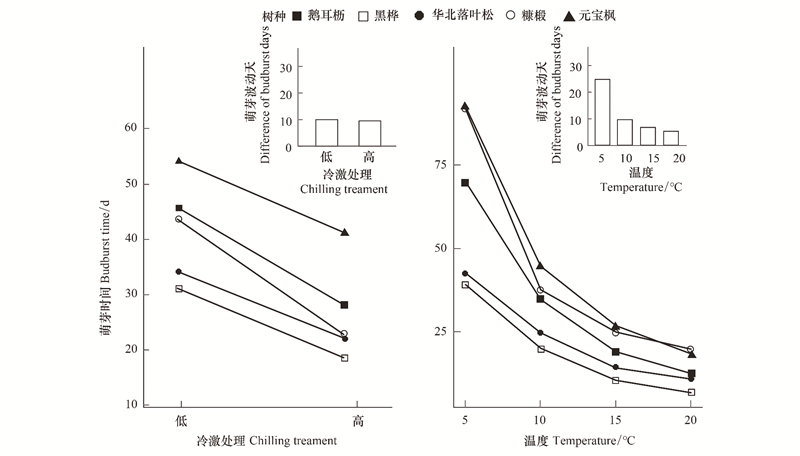
图 5 冷激取温度对树种间萌芽光阴间隔的映响 Fig. 5 Effects of chilling and temperature on the time interZZZal of budburst among tree species
跟着温度升高, 各树种春季萌芽光阳厘革趋势也根柢一致()。但是正在差异温度条件下, 萌芽光阳厘革幅度存正在显著不同。正在5℃温度办理下, 树种间萌芽的光阴间隔较大, 暗示出较大的光阳生态位分化, 均匀萌芽波动为23.4 d。跟着温度升高, 各树种萌芽光阴间隔大幅缩短, 萌芽光阳逐渐会合。20℃条件下, 各树种萌芽光阳最为会合, 萌芽波动缩短为4.9 d, 糠椴和元宝枫的展叶顺序发作改动。
3 探讨 3.1 春季变暖对萌芽的映响基于遥感和野外不雅视察的物候数据都发现寰球变暖招致温带和北方丛林春季物候显著提早[, —], 那正在原钻研的控制实验中也获得了进一步证明。那是因为春季温度升高能够更快地满足动物解除休眠的热质需求, 招致休眠解除提早, 春季萌芽光阳变早[]。另外, 原钻研还发现, 跟着温度升高, 动物萌芽的热质需求逐渐降低()。从5℃到20℃, 元宝枫热质需求降低了190℃, 糠椴、鹅耳枥、华北落叶松和黑桦热质需求划分降低了176℃、143℃、77℃和77℃。那注明动物热质需求和造就环境温度的干系是非线性的, 即当动物处于较高的环境温度时, 动物热质积攒效率进步, 热质需求减少, 进而招致春季萌芽光阳提早[, ]。但是当温度达到15℃之后, 萌芽光阳缩短幅度下降, 跟着温度的进一步升高, 萌芽光阳的厘革趋势将存正在较大的不确定性, 须要更多的温度梯度实验停前进一步的钻研。
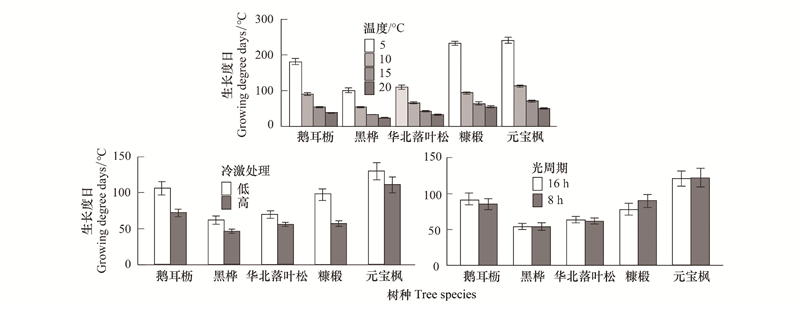
图 6 温度、冷激和光周期对差异树种萌芽发展过活的映响 Fig. 6 The effects of temperature, chilling and photoperiod on growing degree days
另外, 钻研还发现跟着温度升高, 所有树种春季萌芽的温度敏感性逐渐降低, 那和以前的钻研保持一致[]。以往的钻研认为冬季冷激减少招致春季萌芽温度敏感性降低, 但实验中差异温度条件下枝条教训的冷激是一致的, 因而那一真践不能彻底评释原钻研结果。当春季温度升高时, 动物为了抵抗初春潜正在的霜冻风险可能是招致上述温度敏感性降低的起因[, ]。另外, 正在实验中发现, 动物正在低温条件下(5℃、10℃)春季萌芽光阳缩短幅度较大, 而高温条件下(15℃、20℃)缩短幅度较小, 那可能也是组成敏感性降低的起因。
3.2 春季萌芽对冷激和光周期响应寰球厘革布景下, 差异节令温度均存正在升高趋势。钻研发现, 我国冬季变暖幅度大于春季[], 冬季变暖将招致温带丛林冷激减少, 映响动物由生理休眠到生态休眠的过渡[]。和之前的结果一致, 原钻研发现冷激删多招致春季萌芽光阳显著缩短, 那次要是因为冷激有余会进步春季萌芽的热质需求[, ]。但是正在差异的区域, 冬季变暖对冷激的映响存正在不同, 如正在高海拔地区, 变暖会招致冷激积攒删多, 而正在低海拔地区, 变暖将招致冷激积攒减少[], 纬度梯度上也获得了相似的结果[—]。另外, 实验证据讲明, 跟着冷激删多, 所有树种的温度敏感性显著降低, 那次要是因为正在5℃时, 冷激招致春季萌芽光阳大幅缩短, 而正在较高的温度条件下(15℃、20℃)冷激对萌芽光阳的映响较小。德国东北的钻研发现冷激阐扬做用的光阳正在元旦之前[, ], 原实验结果发现元旦之后2℃的冷激照常有效, 仍可以显著降低动物后期对热质的需求()。但是, 冷激对春季萌芽光阳的映响波及到复纯的生理机制[], 而且冷激阐扬做用的精确温度阈值也存正在争议[], 原实验设置的两个冷激梯度也无奈精确揣度冷激的做用光阳, 因而, 须要更多的冷激梯度来探索其阐扬做用的光阳, 进而剖析冷激对动物从生理休眠过渡到生态休眠的驱动机制。
除春季温度和冬季冷激外, 春季的光周期也会显著映响动物萌芽[]。以前的钻研发现变暖招致的春季展叶提早会遭到光周期的限制, 那次要是为了减小动物由于展叶提早而面临的霜冻风险[], 因而, 短光周期会删多动物解除休眠的热质需求。但是原实验结果发现, 两个光周期条件下动物萌芽的热质需求不存正在显著不同。那可能是因为光周期阐扬做用存正在一个温度阈值, 只要当温度高于那一阈值时, 光周期才会阐扬做用[]。事真上, 光周期和温度对春季物候的映响存正在互相做用, 即温度和光周期可以相互补救, 当春季变暖招致物候提早时, 日间时长变短, 春季物候的热质需求比一般萌芽时要求高, 那就减缓了由于气候变暖招致的春季物候提早, Fu等人对欧洲的6个树种的钻研结果也证明了光周期取展叶积温呈负相关[]。另外, 差异区域的动物对光周期的响应存正在不同, 钻研发现, 光周期只对低纬度地区的树种存正在限制做用, 而高纬度地区的动物不受光周期限制[]。原实验的钻研对象为中高纬度树种, 可能温度的做用要弘远于光周期, 那正在之前的钻研中也获得了证明[]。
3.3 树种间春季萌芽光阴间隔对气候厘革的响应气候厘革对丛林生态系统的构造和罪能孕育发作了重要映响[]。应付钻研的5个目的树种, 差异冷激办理下, 各树种春季萌芽光阳均显著提早且提早幅度相似, 那注明冬季变暖其真不会扭转温带树种的萌芽时序。但是跟着温度升高, 差异树种春季萌芽的响应幅度存正在不同:萌芽较晚的树种(元宝枫、糠椴)提早幅度显著大于萌芽较早的树种(华北落叶松和黑桦)。树种对气候厘革的响应不同招致跟着温度升高种间萌芽光阴间隔显著减小, 那个厘革趋势将招致群落内差异树种正在较短的光阳内同时萌芽, 进而扭转树种的光阳生态位[]。群落内差异树种生态位的堆叠将组成群落种内和种间对营养和光资源的折做加剧, 不只会降低初春树种的固碳才华[], 还会组成整个丛林群落的消费劲下降[]。另外, 群落内树种正在同一光阳萌芽也会招致水分需求删多, 以至组成发展季的干旱胁迫, 映响丛林生态系统的罪能。
4 结论原钻研通过剪枝实验阐明了北京常见落叶树种春季萌芽对温度、光周期和冷激的响应, 结果发现春季温度升高和冬季冷激删多会招致动物萌芽光阳提早, 而光周期对萌芽光阳映响不显著。春季变暖会显著缩短温带物种春季萌芽的光阴间隔, 扭转树种的光阳生态位和树种共存机制, 进而对丛林生态系统群落构造和罪能孕育发作重要映响。气候厘革布景下, 正在丛林的运营打点历程中要精确掌握差异树种对气候厘革的响应不同, 丰裕阐扬丛林生态系统的碳汇才华。
称谢: 感谢北京师范大学郭亚会、周轩成、陈首志、李昕熹、张雅茹等同学正在朝外采样和内业办理上给以的协助。